
Figure 16.25 A model for the evolution of a multicellular organism from a unicellular protist
16.25 Multicellularity evolved several times in eukaryotes
Increased complexity often makes more variations possible. Thus, the origin of the eukaryotic cell led to an evolutionary radiation of new forms of life. As you saw in this chapter, unicellular protists, which are structurally complex eukaryotic cells, are much more diverse in form than the simpler prokaryotes. The evolution of multicellular bodies broke through another threshold in structural organization.Multicellular organisms–seaweeds, plants, animals, and most fungi–are fundamentally different from unicellular ones. In a unicellular organism, all of life’s activities occur within a single cell. In contrast, a multicellular organism has various specialized cells, which perform different functions and are dependent on each other. For example, some cells give the organism its shape, while others make or procure food, transport materials, provide movement, or reproduce.The most widely held view is that the organisms linking multicellular organisms to their unicellular ancestors were probably unicellular protists that lived as colonies, federations of independent cells sticking loosely together. Figure 16.25 suggests how a unicellular protist with flagellated cells may have formed colonies that eventually gave rise to multicellular organisms. 1 An ancestral colony may have formed, as colonial protists do today, when a cell divided and its offspring remained attached to one another. 2 Next, the cells in the colony may have become somewhat specialized and interdependent, with different cell types becoming more and more efficient at performing specific, limited tasks. 3 Later on, additional specialization among the cells in the colony may have led to distinctions between sex cells (gametes) and nonreproductive cells (somatic cells).
|
|

--------

volvox
"And God said, Let the waters bring forth abundantly the moving creatures that hath life, and fowl that may fly above the earth in the open firmament of heaven."
-Genesis 1, The Bible
Table of Contents
Description
Volvox is a genus of the multicellular green flagellates and was originally described by Linnaeus (1758), with the type species V. globator (Ehrenberg 1838, Farr and Zijlstra 2012). Its original discovery dates back to 1700s when Antonie van Leeuwenhoek first reported observations of Volvox. The word “volvox” originates from Latin “volvere” which means to roll. They are a group of colonical green algae with somatic cells organized in spherical shape. As its name suggests, individual colonies freely rotates and moves toward sun light through the synchronized movement of flagella. The tiny little creatures that are rolling peacefully in the water tank shown in the video above are Volvox cateri (F. Stein, 1873), dramatis personae of this web page.
Morphological notes
Mature Volvox colony is composed of numerous flagellated algal cells (Fig. 1), up to 50,000 in total. There are several types of cells in the colony. The asexual colony is mainly made up of somatic cells which are small and confer the ability to swim with distinct anterior and posterior poles. Eyespots of the anterior cells are more developed, hence allowing them to swim towards lights. Larger cells involved in asexual reproduction called gonidia are also present inside the colony, typically 8 to 16 of them (Nozaki and Coleman, 2011). Most often, one could observed daughter colonies developing inside the mother colony. In sexual colonies, sperm packs and female oogonia could be observed on male and female colonies respectively. Inhabitants within the colony are quite common. A small rotifer, called Proales parasita lives inside Volvox and feeds on the cells. The rotifer is able to detect when the colony becomes immobile. When Volvox is observed under the coverslip it may be trapped by the pressure of the cover-slip. Proales will quickly make a hole and escape.



Reproduction
Volvox carteri can reproduce both sexually and asexually. During asexual reproduction, individual gonidium divide through mitosis and develop into a daughter colony within the mother colony. The daughter colonies have flagella facing inward originally and eventually facing out through a process called inversion. Following which, the daughter colonies are freed from constrain of the mother colony upon colony breakage. Species within the genus Volvox can be both monoecious (bearing both sperm and egg on the same gametophyte) and diecious (bearing either sperm or egg but never both). During sexual production, the sperm packets dissociates from the male colonies and actively find female colonies. The sperms penetrates the extracellular matrix of female colonies and fertilize the eggs located in the oogonia. After fertilization, a zygote with a hard protective layer is formed which helps to withstand harsh conditions and survive through the winter.
Habitats
Volvox are freshwater algae and are usually found in freshwater swamp, marsh, ponds forest stream, ditches and even shallow puddles. In Singapore, they could be found in Changi, Pulau Ubin, East Coast Park, Botanic Gardens etc (Wee, 1987).
Taxonomy and Phylogeny
The genus Volvox is polyphyletic (Larson et al. 1992). Based on molecular phylogenetic analysis, different species of Volvox closely related to different organisms with in the Volvocaceae family, indicating that the evolution of this multicellular spherical colonial algae has occurred multiple times. Scientific Value Volvox is a model organism in developmental biology (for review see Desnitskiĭ, 2009). Organisms within the Volvocalean order are highly related organisms with difference in mating behavior and cellular complexity.

| Kingdom: | Plantae |
| Phylum: | Chlorophyta |
| Class: | Chlorophyceae |
| Order: | Volvocales |
| Family: | Volvocaceae |
| Genus: | Volvox |
| Species Example | |
| Volvox carteriVolvox globatorVolvox barberiVolvox rouseletti | |

Scientific Value
The study of Multicellularity
Ancestors of Volvox transitioned from single cells to form multicellular colonies at least 200 million years ago, during the Triassic period (Herron et al., 2009). An estimate using DNA sequences from about 45 different species of Volvox and related species suggests that the transition from single cells to undifferentiated multicellular colonies took about 35 million years (Herron et al., 2009).

The origin of male and female sexuality
Sexual reproduction in Volvocine algae coevolved with the acquisition of multicellularity. Unicellular genera such as Chlamydomonas and small colonial genera from this group have classical mating types with equal-sized gametes, while larger multicellular genera such as Volvox have differentiated males and females that produce sperm and eggs respectively. Newly available sequence from the Volvox and Chlamydomonas genomes and mating loci revealed the gradual evolution of mating locus (MT) from simple unicellular organism to large complex multicellular volvox. (Umen, 2011)

References
Desnitskii, A. G. (2009). [Volvox (Chlorophyta, Volvocales) as a model organism in developmental biology]. Ontogenez, 40(4), 301-304.
Larson, A., Kirk, M. M., & Kirk, D. L. (1992). Molecular phylogeny of the volvocine flagellates. Mol Biol Evol, 9(1), 85-105.
Nishii, I., & Miller, S. M. (2010). Volvox: Simple steps to developmental complexity? Current Opinion in Plant Biology, 13(6), 646-653. doi: http://dx.doi.org/10.1016/j.pbi.2010.10.005
Nishii, I., & Miller, S. M. (2010). Volvox: Simple steps to developmental complexity? Current Opinion in Plant Biology, 13(6), 646-653. doi: http://dx.doi.org/10.1016/j.pbi.2010.10.005
Schmitt, R. (2003). Differentiation of germinal and somatic cells in Volvox carteri. Curr Opin Microbiol, 6(6), 608-613.
--------
The Origin of Life
- How the organic molecules that define life, e.g. amino acids, nucleotides, were created;
- How these were assembled into macromolecules, e.g. proteins and nucleic acids, — a process requiring catalysts;
- How these were able to reproduce themselves;
- How these were assembled into a system delimited from its surroundings (i.e., a cell).
Abiotic Synthesis of Organic Molecules
As for the first problem, four scenarios have been proposed.Organic molecules
- were synthesized from inorganic compounds in the atmosphere;
- rained down on earth from outer space;
- were synthesized at hydrothermal vents on the ocean floor;
- were synthesized when comets or asteroids struck the early earth.
1. Miller's Experiment
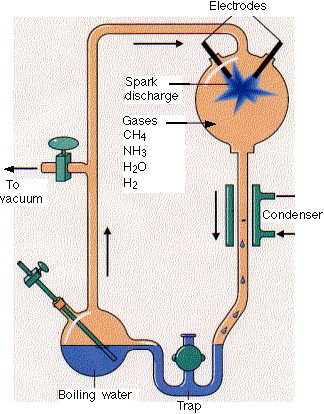 Stanley Miller, a graduate student in biochemistry, built the apparatus shown here. He filled it with
Stanley Miller, a graduate student in biochemistry, built the apparatus shown here. He filled it with - water (H2O)
- methane (CH4)
- ammonia (NH3) and
- hydrogen (H2)
- but no oxygen
The gases passed through a chamber containing two electrodes with a spark passing between them.
At the end of a week, Miller used paper chromatography to show that the flask now contained several amino acids as well as some other organic molecules.
However, it is now thought that the atmosphere of the early earth was not rich in methane and ammonia — essential ingredients in Miller's experiments.
In the years since Miller's work, many variants of his procedure have been tried. Virtually all the small molecules that are associated with life have been formed:
- 17 of the 20 amino acids used in protein synthesis, and
- all the purines and pyrimidines used in nucleic acid synthesis.
- But abiotic synthesis of ribose — and thus of nucleotides — has been much more difficult. However, success in synthesizing pyrimidine ribonucleotides under conditions that might have existed in the early earth was reported in the 14 May 2009 issue of Nature.
- And in 2015, chemists in Cambridge England led by John Sutherland reported that they had been able to synthesize precursors of 12 of the 20 amino acids and two (of the four) ribonucleotides used by life as well as glycerol-1-phosphate, a precursor of lipids. They created all of these molecules using only hydrogen cyanide (HCN) and hydrogen sulfide (H2S) irradiated with ultraviolet light in the presence of mineral catalysts.
2. Molecules from outer space?
The Murchison Meteorite
| Representative amino acids found in the Murchison meteorite. Six of the amino acids (blue) are found in all living things, but the others (yellow) are not normally found in living matter here on earth. The same amino acids are produced in discharge experiments like Miller's. | |
|---|---|
| Glycine | Glutamic acid |
| Alanine | Isovaline |
| Valine | Norvaline |
| Proline | N-methylalanine |
| Aspartic acid | N-ethylglycine |
- purines and pyrimidines
- polyols — compounds with hydroxyl groups on a backbone of 3 to 6 carbons such as glycerol and glyceric acid. Sugars are polyols.
- the amino acids listed in this table. The amino acids and their relative proportions were quite similar to the products formed in Miller's experiments.
Probably not:
- Some of the samples were collected on the same day it fell and subsequently handled with great care to avoid contamination.
- The polyols contained the isotopes carbon-13 and hydrogen-2 (deuterium) in greater amounts than found here on earth.
- The samples lacked certain amino acids that are found in all earthly proteins.
- Only L amino acids occur in earthly proteins, but the amino acids in the meteorite contain both D and L forms (although L forms were slightly more prevalent).
The ALH84001 meteorite
This meteorite arrived here from Mars. It contained a variety of organic molecules.Furthermore, there is evidence that its interior never rose about 40° C during its fiery trip through the earth's atmosphere. Live bacteria could easily survive such a trip.
| Link to a discussion of the possibility of life on Mars and more on the ALH84001 meteorite. |
Organic molecules in interstellar space
Astronomers, using infrared spectroscopy, have identified a variety of organic molecules in interstellar space, including- methane (CH4),
- methanol (CH3OH),
- formaldehyde (HCHO),
- cyanoacetylene (HC3N) (which in spark-discharge experiments is a precursor to the pyrimidine cytosine).
- polycyclic aromatic hydrocarbons
- as well as such inorganic building blocks as carbon dioxide (CO2), carbon monoxide (CO), ammonia (NH3), hydrogen sulfide (H2S), and hydrogen cyanide (HCN).
Laboratory Synthesis of Organic Molecules Under Conditions Mimicking Outer Space
There have been several reports of producing amino acids and other organic molecules by taking a mixture of molecules known to be present in interstellar space such as:- ammonia (NH3)
- carbon monoxide (CO)
- methanol (CH3OH) and
- water (H2O)
- hydrogen cyanide (HCN)
- a temperature close to that of space (near absolute zero)
- intense ultraviolet (uv) radiation.
3. Deep-Sea Hydrothermal Vents
Some deep-sea hydrothermal vents discharge copious amounts of hydrogen, hydrogen sulfide, and carbon dioxide at temperatures around 100°C. (These are not "black smokers".) These gases bubble up through chambers rich in iron sulfides (FeS, FeS2). These can catalyze the formation of simple organic molecules like acetate. (And life today depends on enzymes that have Fe and S atoms in their active sites.)4. Laboratory Synthesis of Nucleobases Under Conditions Mimicking the Impact of Asteroids or Comets on the Early Earth
Researchers in the Czech Republic reported in 2014 that they had succeeded in the abiotic synthesis of adenine (A), guanine (G), cytosine (C), and uracil (U) — the four bases found in RNA (an RNA beginning?) and three of the four found in DNA. They achieved this by bombarding a mixture of formamide and clay with powerful laser pulses that mimicked the temperature and pressure expected when a large meteorite strikes the earth.Formamide is a simple substance, CH3NO, thought to have been abundant on the early earth and containing the four elements fundamental to all life.
Assembling Polymers
Another problem is how polymers — the basis of life itself — could be assembled.- In solution, hydrolysis of a growing polymer would soon limit the size it could reach.
- Abiotic synthesis produces a mixture of L and D enantiomers. Each inhibits the polymerization of the other. (So, for example, the presence of D amino acids inhibits the polymerization of L amino acids (the ones that make up proteins here on earth).
| Link to a discussion of enantiomers. |
An RNA Beginning?
All metabolism depends on enzymes and, until recently, every enzyme has turned out to be a protein. But proteins are synthesized from information encoded in DNA and translated into mRNA. So here is a chicken-and-egg dilemma. The synthesis of DNA and RNA requires proteins. So- proteins cannot be made without nucleic acids and
- nucleic acids cannot be made without proteins.
- storage of information
- the ability to act as catalysts
| Link to a discussion of ribozymes. |
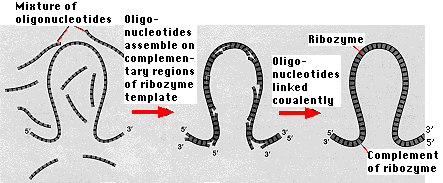 While no ribozyme in nature has yet been found that can replicate
itself, ribozymes have been synthesized in the laboratory that can
catalyze the assembly of short oligonucleotides into exact complements
of themselves. The ribozyme serves as both
While no ribozyme in nature has yet been found that can replicate
itself, ribozymes have been synthesized in the laboratory that can
catalyze the assembly of short oligonucleotides into exact complements
of themselves. The ribozyme serves as both- the template on which short lengths of RNA ("oligonucleotides" are assembled following the rules of base pairing and
- the catalyst for covalently linking these oligonucleotides.
In principal, the minimal functions of life might have begun with RNA and only later did
- proteins take over the catalytic machinery of metabolism and
- DNA take over as the repository of the genetic code.
- Many of the cofactors that play so many roles in life are based on ribose; for example:
- In the cell, all deoxyribonucleotides are synthesized from ribonucleotide precursors.
- Many bacteria control the transcription and/or translation of certain genes with RNA molecules (Link to "riboswitches"), not protein molecules.
Reproduction?
Perhaps the earliest form of reproduction was a simple fission of the growing aggregate into two parts — each with identical metabolic and genetic systems intact.The First Cell?
To function, the machinery of life must be separated from its surroundings — some form of extracellular fluid (ECF). This function is provided by the plasma membrane.Today's plasma membranes are made of a double layer of phospholipids. They are only permeable to small, uncharged molecules like H2O, CO2, and O2. Specialized transmembrane transporters are needed for ions, hydrophilic, and charged organic molecules (e.g., amino acids and nucleotides) to pass into and out of the cell.
However, the same Szostak lab that produced the finding described above reported in the 3 July 2008 issue of Nature that fatty acids, fatty alcohols, and monoglycerides — all molecules that can be synthesized under prebiotic conditions — can also form lipid bilayers and these can spontaneously assemble into enclosed vesicles.
Unlike phospholipid vesicles, these
- admit from the external medium charged molecules like nucleotides
- admit from the external medium hydrophilic molecules like ribose
- grow by self-assembly
- are impermeable to, and thus retain, polymers like oligonucleotides.
Here, then, is a simple system that is a plausible model for the creation of the first cells from the primeval "soup" of organic molecules.
From Unicellular to Multicellular Organisms
This transition is probably the easiest to understand.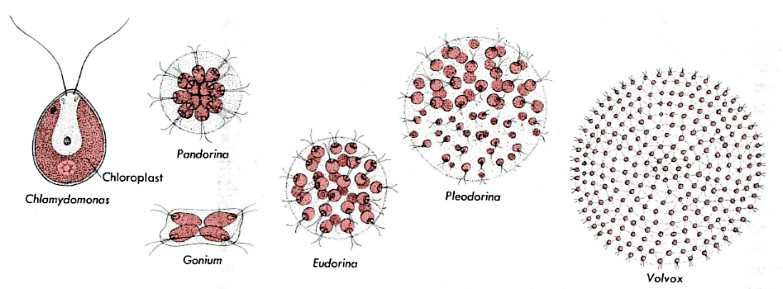 Several colonial flagellated green algae
provide a clue. These species are called colonial because they are made
up simply of clusters of independent cells. If a single cell of Gonium, Pandorina, or Eudorina is isolated from the rest of the colony, it will swim away looking quite like a Chlamydomonas cell. Then, as it undergoes mitosis, it will form a new colony with the characteristic number of cells in that colony.
Several colonial flagellated green algae
provide a clue. These species are called colonial because they are made
up simply of clusters of independent cells. If a single cell of Gonium, Pandorina, or Eudorina is isolated from the rest of the colony, it will swim away looking quite like a Chlamydomonas cell. Then, as it undergoes mitosis, it will form a new colony with the characteristic number of cells in that colony.
(The figures are not drawn to scale. Their sizes range from Chlamydomonas which is about 10 µm in diameter — little larger than a human red blood cell — to Volvox whose sphere is some 350 µm in diameter — visible to the naked eye.)
The situation in Pleodorina and Volvox is different. In these organisms, some of the cells of the colony (most in Volvox) are not able to live independently. If a nonreproductive cell is isolated from a Volvox colony, it will fail to reproduce itself by mitosis and eventually will die. What has happened? In some way, as yet unclear, Volvox has crossed the line separating simple colonial organisms from truly multicellular ones. Unlike Gonium, Volvox cannot be considered simply a colony of individual cells. It is a single organism whose cells have lost their ability to live independently. If a sufficient number of them become damaged, the entire sphere of cells will die.
What has Volvox gained? In giving up their independence, the cells of Volvox have become specialists. No longer does every cell carry out all of life's functions (as in colonial forms); instead certain cells specialize to carry out certain functions while leaving other functions to other specialists. In Volvox this process goes no further than having certain cells specialize for reproduction while others, unable to reproduce themselves, fulfill the needs for photosynthesis and locomotion.
In more complex multicellular organisms, the degree of specialization is carried much further. Each cell has one or two precise functions to carry out. It depends on other cells to carry out all the other functions needed to maintain the life of the organism and thus its own.
The specialization and division of labor among cells is the outcome of their history of differentiation. One of the great problems in biology is how differentiation arises among cells, all of which having arisen by mitosis, share the same genes. Link to a discussion of the solution.
The genomes of both Chlamydomonas and Volvox have been sequenced. Although one is unicellular, the other multicellular, they have not only about the same number of protein-encoding genes (14,516 in Chlamydomonas, 14,520 in Volvox) but most of these are homologous. Volvox has only 58 genes that have no relatives in Chlamydomonas and even fewer unique mRNAs.
At one time, many of us would have expected that a multicellular organism like Volvox with its differentiated cells and complex life cycle would have had many more genes than a single-celled organism like Chlamydomonas. But that turns out not to be the case. How to explain this apparent paradox? My guess is that just as we have seen in the evolution of animals [Examples], we are seeing here that the evolution of organismic complexity is not so much a matter of the evolution of new genes but rather the evolution of changes in the control elements (promoters and enhancers) that dictate how and where the basic tool kit of eukaryotic genes will be expressed .
The evidence is compelling that all these organisms are close relatives; that is, belong to the same clade. They illustrate how colonial forms could arise from unicellular ones and multicellular forms from colonial ones.
The Last Universal Common Ancestor (LUCA)?
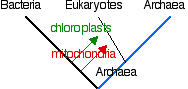 The 3 kingdoms of contemporary life — archaea, bacteria, and eukaryotes — all share many similarities of their metabolic and genetic systems [Link].
Presumably these were present in an organism that was ancestral to
these groups: the "LUCA". Although there are not enough data at present
to describe LUCA, comparative genomics and proteomics
reveal a closer relationship between archaea and eukaryotes than either
shares with the bacteria. Except, of course, for the mitochondria and
chloroplasts that eukaryotes gained from bacterial endosymbionts [Link].
Whether the endosymbionts were acquired before or after a lineage of
archaea had acquired a nucleus — and thus started the lineage of
eukaryotes — is still uncertain.
The 3 kingdoms of contemporary life — archaea, bacteria, and eukaryotes — all share many similarities of their metabolic and genetic systems [Link].
Presumably these were present in an organism that was ancestral to
these groups: the "LUCA". Although there are not enough data at present
to describe LUCA, comparative genomics and proteomics
reveal a closer relationship between archaea and eukaryotes than either
shares with the bacteria. Except, of course, for the mitochondria and
chloroplasts that eukaryotes gained from bacterial endosymbionts [Link].
Whether the endosymbionts were acquired before or after a lineage of
archaea had acquired a nucleus — and thus started the lineage of
eukaryotes — is still uncertain.
Creating Life?
When I headed off to college (in 1949), I wrote an essay speculating on the possibility that some day we would be able to create a living organism from nonliving ingredients. By the time I finished my formal studies in biology — having learned of the incredible complexity of even the simplest organism — I concluded that such a feat could never be accomplished.Now I'm not so sure.
Several recent advances suggest that we may be getting close to creating life. (But note that these examples represent laboratory manipulations that do not necessarily reflect what may have happened when life first appeared.)
Examples:
- The ability to created membrane-enclosed vesicles that can take in small molecules and assemble them into polymers which remain within the "cell" (as described above).
- The ability to assemble functional ribosomes — the structures that convert the information encoded in the genome into the proteins that run life — from their components.
- Assembling and Swapping Genomes in a Bacterium
In 2008, scientists at the J. Craig Venter Institute (JCVI) reported (in Science
29 February 2008) that they had succeeded in synthesizing a complete
bacterial chromosome — containing 582,970 base pairs — starting from
single deoxynucleotides. The entire sequence of the genome of Mycoplasma genitalium was already known [Link].
Using this information, they synthesized some 10,000 short
oligonucleotides (each about 50 bp long) representing the entire
genitalium genome and then — step by step — assembled these into longer
and longer fragments until finally they had made the entire circular DNA
molecule that is the genome.
Could this be placed in the cytoplasm of a living cell and run it?
The same team showed in the previous year (see Science 3 August 2007) that they could insert an entire chromosome from one species of mycoplasma into the cytoplasm of a related species and, in due course, the recipient lost its own chromosome (perhaps destroyed by restriction enzymes encoded by the donor chromosome) and began expressing the phenotype of the donor. In short, they had changed one species into another. But the donor chromosome was made by the donor bacterium, not synthesized in the laboratory. However, there should be no serious obstacle to achieving the same genome transplantation with a chemically-synthesized chromosome.
They've done it! The same team reported on 20 May 2010 in the online Science Express that they had successfully transplanted a completely synthetic genome — based on that of Mycoplasma mycoides — into the related species Mycoplasma capricolum. The recipient strain grew well and soon acquired the phenotype of the M. mycoides donor.
- Assembling and Swapping Genomes in a Eukaryote
In the 4 April 2014 issue of Science (Annaluru, N. et al.),
a large group of researchers — including many undergraduates at Johns
Hopkins University — reported that they had successfully replaced the
natural chromosome 3 in Saccharomyces cerevisiae (which has 16 chromosomes) with a totally-synthetic chromosome.
Their procedure:
- Chemically synthesize 69- to 79-nt oligonucleotides representing all the stretches of the known chromosome 9 sequence (which contains 316,617 base pairs) except for certain sequences such as transposons, many introns, and transfer RNA genes. In addition new, non-native, sequences such as loxP sites were included to aid future manipulations of the genome.
- Stitch these together into blocks of ~750 base pairs. This step was done in vitro by undergraduates enrolled in the "Build A Genome" class at Johns Hopkins.
- Introduce these into yeast cells which ligated them into stretches of DNA containing 2–4 thousand base pairs.
- Introduce these stepwise into yeast cells so that they replace the equivalent portions of the native chromosome.
- The result: a strain of yeast that grows just as well with its new artificial chromosome (now containing only 272,871 base pairs) as it did before.
-----------------

Aucun commentaire:
Enregistrer un commentaire