Embryogenèse somatique
L'embryogenèse somatique est une technique couramment utilisée en culture in vitro. Elle permet de générer un embryon à partir d'un cal ou de suspensions cellulaires. Cette technique fait appel à la notion de totipotence (spécifique des végétaux), en effet on part ici de cellules somatiques et non de cellules germinales.
Sommaire
Principe
L'embryogenèse somatique permet d'obtenir des plantules génétiquement identiques à la plante mère. Dans une graine, on trouve la future plante sous forme d'embryon (embryon zygotique) qui résulte de la reproduction sexuée. Cette technique consiste alors à provoquer l'apparition des tissus végétaux mis en culture in vitro, qui provoque de nombreuses divisions cellulaires. Cette embryogenèse somatique génère alors des embryons dans ces divisions cellulaires ou dans les cals, c'est-à-dire un amas de cellules indifférenciées (qui ont été dédifférenciées sur l'explant de la plante mère avec le phénomène de totipotence végétale).Sous certaines conditions, les cultures cellulaires s'organisent ensuite en nombreux petits massifs à structure bipolaire (avec un méristème de racines et un méristème de tiges) nommés embryons somatiques. Comme les embryons zygotiques présents dans les graines, les embryons somatiques obtenus à partir de cellules non sexuées (sans fécondation) se développent en un nombre illimité de plantes génétiquement identiques. C'est actuellement la technique la plus performante pour la multiplication végétative des conifères.
Les principales étapes
Étape 1
On induit de nombreuses divisions cellulaires à partir des tissus mis en culture. Le milieu est complémenté par de fortes doses d'auxine (2,4D) et de cytokinines. Le 2,4D est une auxine de synthèse. Elle est perçue comme une auxine par la plante mais n'est pas dégradée. Son accumulation provoque alors un choc auxinique : les cellules prolifèrent et forment un cal cellulaire.Étape 2
Le cal est transféré sur un milieu d'induction de l'embryogenèse. Ce milieu est seulement complémenté en 2,4D. Le cal cellulaire devient alors embryogène. On peut ainsi découper ce cal en plusieurs morceaux pour augmenter la taille de la population végétale.Étape 3
Le cal est transféré sur un milieu d'expression ou milieu banc. Ce milieu ne contient aucune hormone et va permettre le développement des embryons (que le 2,4D inhibait).Étape 4
Les embryons vont ensuite maturer (au bout de 5 semaines), puis germer. On obtient des plantules somatiques au bout de 12 semaines, elles vont alors pouvoir être cultivées en sol pour finir leur croissance.| Composés | Milieu blanc | Milieu d'induction | Milieu d'expression |
|---|---|---|---|
| Macroéléments de Murashige & skoog | 200 ml | 200 ml | 200ml |
| FeEDTA | 10 ml | 10 ml | 10 ml |
| Oligo-éléments | 1 ml | 1 ml | 1 ml |
| Vitamines | 1 ml | 1 ml | 1 ml |
| Saccharose | 30 ml | 30 ml | 30 ml |
| Gélose | 10 ml | 10 ml | 10 ml |
| 2,4D | 0 | 10-6M | 10-6M |
| Cytokinine | 0 | 10-6M | 0 |
Schéma récapitulatif
Applications
- Obtention rapide de semences d'élites pour les espèces ligneuses.
- Obtention de semences pour des variétés stériles (pommes de terre polyploïdes, bananier...).
- Possibilité de culture en réacteur : production de semences artificielles à grande échelle (gymnospermes).
- Obtention de germplasme, utilisé ainsi lors de la création de banques de souches pour la conservation génétique des plantes.
- Sauvegarde et multiplication de plantes en voie d'extinction.
Voir aussi
Articles connexes
Liens externes
- Exemple du pin et de l'épinette blanche
- La multiplication d'arbres
- Régénération via l'organogenèse ou l'embryogenèse somatique
- Vitrovariation et régénération par embryogenèse somatique à partir d'embryons mûrs de blé tendre
- Inoplant
Somatic embryogenesis
From Wikipedia, the free encyclopedia
| This article needs additional citations for verification. (February 2015) (Learn how and when to remove this template message) |
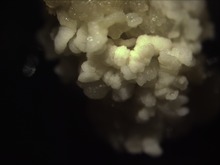
Switchgrass somatic embryos
Contents
- 1 Direct and indirect embryogenesis
- 2 Plant regeneration via somatic embryogenesis
- 3 Factors influencing somatic embryogenesis
- 4 Uses of somatic embryogenesis
- 5 Problems associated with somatic embryogenesis
- 6 Tracking and fate maps
- 7 Angiosperms
- 8 Gymnosperms
- 9 See also
- 10 References
- 11 External links
Direct and indirect embryogenesis
Somatic embryogenesis has been described to occur in two ways: directly or indirectly.[6] Direct embryogenesis occurs when embryos are started directly from explant tissue creating an identical clone. Indirect embryogenesis occurs when explants produced undifferentiated, or partially differentiated, cells (often referred to as callus) which then is maintained or differentiated into plant tissues such as leaf, stem, or roots.Plant regeneration via somatic embryogenesis
Plant regeneration via somatic embryogenesis occurs in five steps: initiation of embryogenic cultures, proliferation of embryogenic cultures, prematuration of somatic embryos, maturation of somatic embryos and plant development on nonspecific media. Initiation and proliferation occur on a medium rich in auxin, which induces differentiation of localized meristematic cells. The auxin typically used is 2,4-D. Once transferred to a medium with low or no auxin, these cells can then develop into mature embryos. Germination of the somatic embryo can only occur when it is mature enough to have functional root and shoot apices [2]Factors influencing somatic embryogenesis
Factors and mechanisms controlling cell differentiation in somatic embryos are relatively ambiguous. Certain compounds excreted by plant tissue cultures and found in culture media have been shown necessary to coordinate cell division and morphological changes.[7] These compounds have been identified by Chung et al.[8] as various polysaccharides, amino acids, growth regulators, vitamins, low molecular weight compounds and polypeptides. Several signaling molecules known to influence or control the formation of somatic embryos have been found and include extracellular proteins, arabinogalactan proteins and lipochitooligosaccharides. Temperature and lighting can also affect the maturation of the somatic embryo.Uses of somatic embryogenesis
- Plant transformations
- Mass propagation[9]
The development of somatic embryogenesis procedures has given rise to research on seed storage proteins (SSPs) of woody plants for tree species of commercial importance, i.e., mainly gymnosperms, including white spruce. In this area of study, SSPs are used as markers to determine the embryogenic potential and competency of the embryogenic system to produce a somatic embryo biochemically similar to its zygotic counterpart (Flinn et al. 1991, Beardmore et al. 1997).[10][11]
Grossnickle et al. (1992)[12] compared interior spruce seedlings with emblings during nursery development and through a stock quality assessment program immediately before field outplanting. Seedling shoot height, root collar diameter, and dry weight increased at a greater rate in seedlings than in emblings during the first half of the first growing season, but thereafter shoot growth was similar among all plants. By the end of the growing season, seedlings were 70% taller than emblings, had greater root collar diameter, and greater shoot dry weight. Root dry weight increased more rapidly in seedlings than in emblings during the early growing season
During fall acclimation, the pattern of increasing dormancy release index and increasing tolerance to freezing was similar in both seedlings and emblings. Root growth capacity decreased then increased during fall acclimation, with the increase being greater in seedlings.
Assessment of stock quality just prior to planting showed that: emblings had greater water use efficiency with decreasing predawn shoot water potential compared with seedlings; seedlings and emblings had similar water movement capability at both high and low root temperatures; net photosynthesis and needle conductance at low root temperatures were greater in seedlings than in emblings; and seedlings had greater root growth than emblings at 22°C root, but root growth among all plants was low at 7.5°C root temperature.
Growth and survival of interior spruce 313B Styroblock® seedlings and emblings after outplanting on a reforestation site were determined by Grossnickle and Major (1992).[13] For both seedlings and emblings, osmotic potential at saturation (ψsat) and turgor loss point (ψtip) increased from a low of -1.82 and -2.22 MPa, respectively, just prior to planting to a seasonal high of -1.09 and -1.21 MPa, respectively, during active shoot elongation. Thereafter, seedlings and emblings (ψsat) and (ψtip) declined to -2.00 and -2.45 MPa, respectively, at the end of the growing season, which coincided with the steady decline in site temperatures and a cessation of height growth. In general, seedlings and emblings had similar ψsat and ψtip values through the growing season, and also had similar shifts in seasonal patterns of maximum modulus of elasticity, sympalstic fraction, and relative water content at turgor loss point.
Grossnickle and Major (1992)[13] found that year-old and current-year needles of both seedlings and emblings had a similar decline in needle conductance with increasing vapour pressure deficit. Response surface models of current-year needles net photosynthesis (Pn) response to vapour pressure deficit (VPD) and photosynthetically active radiation (PAR) showed that emblings had 15% greater Pn at VPD of less than 3.0 kPa and PAR greater than 1000 μmol m−2s−1. Year-old and current-year needles of seedlings and emblings showed similar patterns of water use efficiency.
Rates of shoot growth in seedlings and emblings through the growing season were also similar to one another. Seedlings had larger shoot systems both at the time of planting and at the end of the growing season. Seedlings also had greater root development than emblings through the growing season, but root:shoot ratios for the 2 stock types were similar at the end of the growing season, when the survival rates for seedlings and emblings were 96% and 99%, respectively.
Problems associated with somatic embryogenesis
| This section requires expansion. (February 2015) |
- High chance of mutations
- Difficult method
- Loss of regenerative ability
- High percentage of albino shoots during regeneration
- Not possible with all plant species and must be optimized for each species and its use
Tracking and fate maps
Understanding the formation of a somatic embryo through establishment of morphological and molecular markers is important for construction of a fate map. The fate map is the foundation in which to build further research and experimentation. Two methods exist to construct a fate map: synchronous cell-division and time-lapse tracking. The latter typically works more consistently because of cell-cycle-altering chemicals and centrifuging involved in synchronous cell-division.[14]Angiosperms
Embryo development in angiosperms is divided into several steps. The zygote is divided asymmetrically forming a small apical cell and large basal cell. The organizational pattern is formed in the globular stage and the embryo then transitions to the cotyledonary stage.[15] Embryo development differs in monocots and dicots. Dicots pass through the globular, heart-shaped, and torpedo stages while monocots pass through globular, scuetellar, and coleoptilar stages.[16]Many culture systems induce and maintain somatic embryogenesis by continuous exposure to 2,4-dichlorophenoxyacetic acid. Abscisic acid has been reported to induce somatic embryogenesis in seedlings. After callus formation, culturing on a low auxin or hormone free media will promote somatic embryo growth and root formation. In monocots, embryogenic capability is usually restricted to tissues with embryogenic or meristematic origin. Somatic cells of monocots differentiate quickly and then lose mitotic and morphogenic capability. Differences of auxin sensitivity in embryogenic callus growth between different genotypes of the same species show how variable auxin responses can be.[17]
Carrot Daucus carota was the first and most understood species with regard to developmental pathways and molecular mechanisms.[14] Time-lapse tracking by Toonen et al. (1994) showed that morphology of competent cells can vary based on shape and cytoplasm density. Five types of cells were identified from embryonic suspension: spherical cytoplasm-rich, spherical vacuolated, oval vacuolated, elongated vacuolated, and irregular shaped cells. Each type of cell multiplied with certain geometric symmetry. They developed into symmetrical, asymmetrical, and aberrantly-shaped cell clusters that eventually formed embryos at different frequencies.[18] This indicates that organized growth polarity do not always exist in somatic embryogenesis.[14]
Aucun commentaire:
Enregistrer un commentaire